Hey guys, here is the link to my podcast on Ghrelin. :) Below is the transcript. ENJOY! :)
Welcome to the show! Thank you for tuning in Flash in the Pan, and I’m your host Linfei. For today’s episode, we will be talking about why we are always hungry?! Have you ever felt like; umm I want to eat something. Despite you just had a delicious meal, and you were full five minutes ago, our stomachs keep on growling and telling us: FOOD!! We want more! Well, this is because Ghrelin is stimulating the brain and giving rise to an increase in appetite. Ghrelin is the reason why we love food so much and we can’t help but eat more. So, what is this foodie, Ghrelin? Ghrelin is a water-soluble, 28-amino acid hunger stimulating peptide and hormone that is responsible for not only the stimulation in the brain (in order to increase in appetite), but also the accumulation of lipids in visceral fatty tissue. According to the National Digestive Diseases Clearing House, Ghrelin is produced in the stomach and upper intestine in the absence of food in the digestive system. It is produced mainly by P/D1 cells lining the fundus of the human stomachs and epsilon cells of the pancreas that stimulates appetite. Ghrelin is also produced in the hypothalamic arcuate nucleus, where it stimulates the secretion of growth hormone from the anterior pituitary gland. Also, Receptors for ghrelin are expressed by neurons in the arcuate nucleus and the lateral hypothalamus. The ghrelin receptor is a G protein-coupled receptor, formerly known as the GHS receptor (growth hormone secretagogue receptor).
On the other hand, Ghrelin can also be secreted by the lungs, pancreatic islets, gonads, adrenal cortex, placenta, kidney, and brain. The diversity in areas of ghrelin production indicates that this hormone has various and numerous biological functions, such as gastrointestinal tract, learning and memory, anxiety response and depression, sleep duration, appetite inducer, chronic stress and PTSD and body weight regulation. So, how does ghrelin work?
Ghrelin has a negative feedback loop that ghrelin is involved in regulating our bodies. As the body loses weight, ghrelin levels rise in response to the energy deficit. Studies had researched this topic through fasting, but an experiment had not yet been run. Studies found that ghrelin levels increased in response to modest weight loss resulting from exercise without a reduction in food intake. Thus, ghrelin also plays a role in the body’s adaptive response to weight loss, either through reducing food intake or increasing levels of physical activity. Ghrelin and synthetic ghrelin mimetic (the growth hormone secretagogues) increase food intake and increase fat mass by an action exerted at the level of the hypothalamus. They activate cells in the arcuate nucleus that include the orexigenic neuropeptide Y (NPY) neurons. Ghrelin also activates the mesolimbic cholinergic-dopaminergic reward link, a circuit that communicates the hedonic and reinforcing aspects of natural rewards, such as food.
Do not underestimate ghrelin! It is not just a foodie. It is also a harm/threat to our health. As mentioned before, ghrelin also favors the accumulation of lipids in visceral fatty tissue. This type of accumulated fat in the region of the abdomen that is deemed to be most harmful, as it is accompanied by comorbilities. Also, visceral obesity is related to the risk of getting higher blood pressure or type II diabetes. Moreover, being located in the abdominal zone and in direct contact with the liver, this type of fatty tissue favours the formation of liver fat and increases the risk of developing resistance to insulin. Nevertheless, there are ways to regulate ghrelin from releasing. Factors could affect ghrelin levels are nutrient stimulation of the gastrointestinal tract, diet composition and weight loss. Moreover, as all hormones work on a feed back loop, something triggers its release and something stops it from being released. Therefore, another hormone that is found to be secreted from the stomach in response to food ingestion could moderate ghrelin levels. This hormone is peptide P YY3-36. It can blunt ghrelin secretions; thus, it creates lower appetite and reduces food intake. Leptin could regulate ghrelin as well. Leptin is a 16-kDa adipokine that plays a vital role in regulating energy intake and expenditure, including appetite and hunger, metabolism and behavior. It is one of the most important adipose-derived hormones. Leptin functions by binding to the Leptin receptor (LEP-R), and it is located on chromosome 7 in humans. Besides hormone, scientists have developed an anti-obesity vaccine to regulate ghrelin. The vaccine uses the immune system, specifically antibodies, to bind to selected targets, directing the body's own immune response against them. This prevents ghrelin from reaching the central nervous system, thus producing a desired reduction in weight gain.
Best way to control weight and stay healthy is to eat slowly and to wait twenty minutes to let the body signal to you that you are full. That’s all for today! Thank you for listening, and remember to give your stomach a twenty-minute-break! See you next time!

Work Cited:
1. Alvarez-Castro, P., et al. “Marked GH Secretion after Ghrelin Alone or Combined with GH Releasing Hormone (GHRH) in Obese Patients.” Clinical Endocrinology, 61 (2), 2004. 250-255. Retrieved from: http://www.userwebs.pomona.edu/~ejc14747/student%20presentations/McKibben_Ghrelin.pdf
2. Basque Research. (2009, May 26). Action Of Ghrelin Hormone Increases Appetite And Favors Accumulation Of Abdominal Fat. ScienceDaily. Retrieved February 24, 2014 from www.sciencedaily.com/releases/2009/05/090520055519.htm
3. Kojima, Masayasu, and Kenji Kangawa. “Ghrelin: Structure and Function.” Physiological Reviews, 85 (2), 2005. 495-522. Retrieved from: http://www.userwebs.pomona.edu/~ejc14747/student%20presentations/McKibben_Ghrelin.pdf
4. Williams, D. L., & Cummings, D. E. (2005). Regulation of ghrelin in physiologic and pathophysiologic states. Informally published manuscript, The American Society for Nutritional Sciences, Retrieved from http://nutrition.highwire.org/content/135/5/1320.full
5. Wren, A.M., et al. “Ghrelin Enhances Appetite and Increases Food Intake in Humans.” Journal of Clinical Endocrinology and Metabolism, 86 (12), 2001. Retrieved from: http://www.userwebs.pomona.edu/~ejc14747/student%20presentations/McKibben_Ghrelin.pdf
6. http://www.news-medical.net/health/Ghrelin-What-is-Ghrelin.aspx
7. http://www.healthaliciousness.com/blog/The-Hormones-of-Digestion.php
*Colored parts are in the actual audio
Do not underestimate ghrelin! It is not just a foodie. It is also a harm/threat to our health. As mentioned before, ghrelin also favors the accumulation of lipids in visceral fatty tissue. This type of accumulated fat in the region of the abdomen that is deemed to be most harmful, as it is accompanied by comorbilities. Also, visceral obesity is related to the risk of getting higher blood pressure or type II diabetes. Moreover, being located in the abdominal zone and in direct contact with the liver, this type of fatty tissue favours the formation of liver fat and increases the risk of developing resistance to insulin. Nevertheless, there are ways to regulate ghrelin from releasing. Factors could affect ghrelin levels are nutrient stimulation of the gastrointestinal tract, diet composition and weight loss. Moreover, as all hormones work on a feed back loop, something triggers its release and something stops it from being released. Therefore, another hormone that is found to be secreted from the stomach in response to food ingestion could moderate ghrelin levels. This hormone is peptide P YY3-36. It can blunt ghrelin secretions; thus, it creates lower appetite and reduces food intake. Leptin could regulate ghrelin as well. Leptin is a 16-kDa adipokine that plays a vital role in regulating energy intake and expenditure, including appetite and hunger, metabolism and behavior. It is one of the most important adipose-derived hormones. Leptin functions by binding to the Leptin receptor (LEP-R), and it is located on chromosome 7 in humans. Besides hormone, scientists have developed an anti-obesity vaccine to regulate ghrelin. The vaccine uses the immune system, specifically antibodies, to bind to selected targets, directing the body's own immune response against them. This prevents ghrelin from reaching the central nervous system, thus producing a desired reduction in weight gain.
Best way to control weight and stay healthy is to eat slowly and to wait twenty minutes to let the body signal to you that you are full. That’s all for today! Thank you for listening, and remember to give your stomach a twenty-minute-break! See you next time!
 |
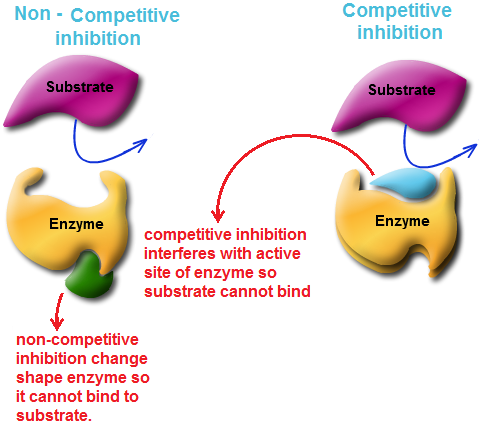
| Three types of Chemical Pathway |
 |
| Leptin's sturcture |
 |
| Ghrelin's sturcture |
 |
| This is how Ghrelin works! |
 |
| Ghrelin action to stimulate GH secretion |
 |
| Regulation |
 |
| Watch out Ghrelin!! |
Work Cited:
1. Alvarez-Castro, P., et al. “Marked GH Secretion after Ghrelin Alone or Combined with GH Releasing Hormone (GHRH) in Obese Patients.” Clinical Endocrinology, 61 (2), 2004. 250-255. Retrieved from: http://www.userwebs.pomona.edu/~ejc14747/student%20presentations/McKibben_Ghrelin.pdf
2. Basque Research. (2009, May 26). Action Of Ghrelin Hormone Increases Appetite And Favors Accumulation Of Abdominal Fat. ScienceDaily. Retrieved February 24, 2014 from www.sciencedaily.com/releases/2009/05/090520055519.htm
3. Kojima, Masayasu, and Kenji Kangawa. “Ghrelin: Structure and Function.” Physiological Reviews, 85 (2), 2005. 495-522. Retrieved from: http://www.userwebs.pomona.edu/~ejc14747/student%20presentations/McKibben_Ghrelin.pdf
4. Williams, D. L., & Cummings, D. E. (2005). Regulation of ghrelin in physiologic and pathophysiologic states. Informally published manuscript, The American Society for Nutritional Sciences, Retrieved from http://nutrition.highwire.org/content/135/5/1320.full
5. Wren, A.M., et al. “Ghrelin Enhances Appetite and Increases Food Intake in Humans.” Journal of Clinical Endocrinology and Metabolism, 86 (12), 2001. Retrieved from: http://www.userwebs.pomona.edu/~ejc14747/student%20presentations/McKibben_Ghrelin.pdf
6. http://www.news-medical.net/health/Ghrelin-What-is-Ghrelin.aspx
7. http://www.healthaliciousness.com/blog/The-Hormones-of-Digestion.php
*Colored parts are in the actual audio